





















پیام رسانی سلولی
لطفا برای بهتر شدن شرایط داروسازی کشور و بهبود سلامت عمومی ایران در نظر سنجی زیر شرکت کنید.
برای شرکت در نظر سنجی کلیک کنید.
سلول ها از طریق ارسال و دریافت سیگنال ها با هم ارتباط برقرار می کنند.
سیگنال ها از محیط و یا از یک سلول دیگر می آیند. برای آغاز یک واکنش، این سیگنال ها باید در سراسر غشاء انتقال پیدا کنند. گاهی اوقات سیگنال به تنهایی می تواند از غشاء عبور کند. در دیگر موارد از طریق واکنش با پروتئین های گیرنده که با هر دو طرف داخل و خارج سلول در ارتباط است عمل می کند. در این مورد، تنها سلول هایی که گیرنده های صحیحی در سطوح خود دارند می توانند به سیگنال پاسخ بدهند.

گیرنده های سلولی:
- گیرنده های تیروزین کینازی (RTK):
گیرنده های تیروزین کینازی (RTK) شامل گیرنده های انسولین و برخی از فاکتورهای رشد مانند فاکتور رشد اپیدرمی (EGF) و فاکتور رشد مشتق از پلاکت (PDGF) می باشند. گیرنده های تیروزین کینازی از نوع پروتئین های غشایی بوده و هر گیرنده به صورت یک دیمر است که هر منومر آن دارای دو بخش آلفا و بتا می باشد. گیرنده انسولین به صورت تترامر a2B2 است. برخلاف انسولین، دو منومر گیرنده های دیگر به صورت منومرهای مجزا بوده و با فعال شدن ایجاد دیمر می کنند.

اتصال فاکتور رشد به ناحیه خارج سلولی RTK، موجب دیمری شدن زیر واحدهای RTK شده که خود منجر به فعال شدن ناحیه سیتوپلاسمی دارای فعالیت کینازی می شود. بخش سیتوزولی گیرنده توسط کیناز در چندین زیر واحد تیروزین فسفریله می شود (اتوفسفریلاسیون). سپس هر زیر واحد تیروزین فسفریله به عنوان جایگاه تشخیص یا اتصال برای سایر پروتئین هایی که سوبسترای RTK هستند عمل می کند.
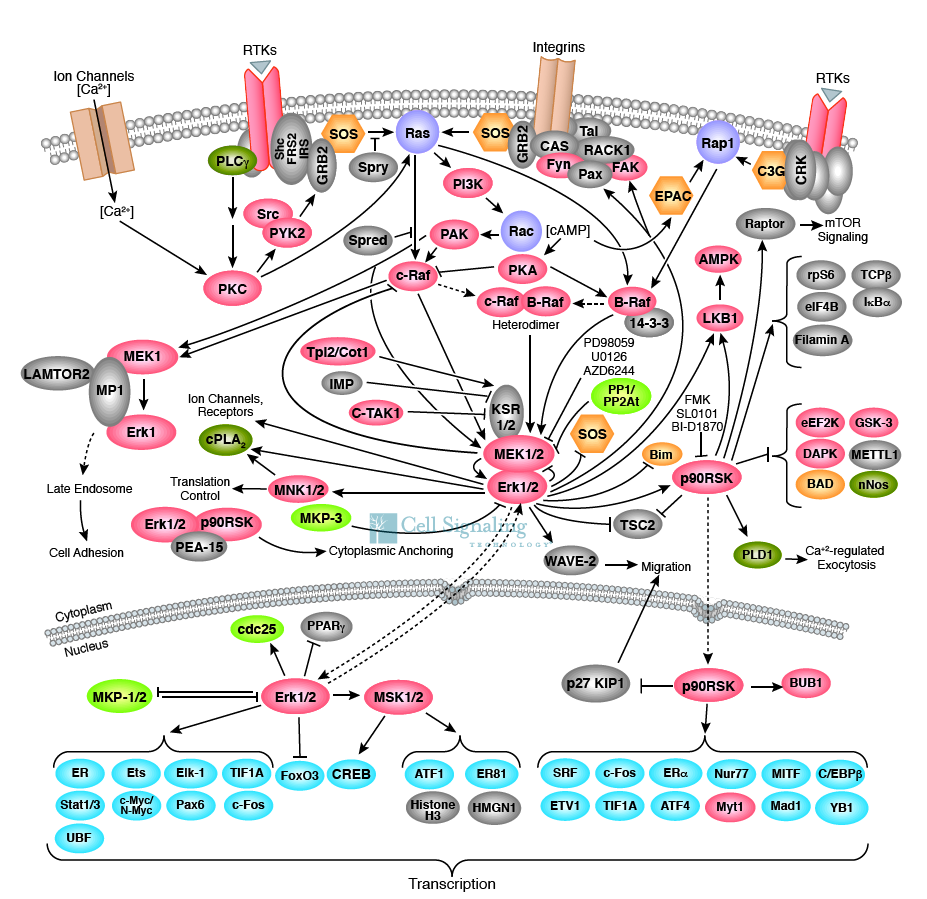
پروتئین Ras یکGTPase کوچک منومریک متصل شونده بهGTP است که یک تنظیم کننده حیاتی در تکثیر سلول می باشد. حدود 30% کل تومورهای انسانی دارای سلول های بیان کننده انکوژن های Ras جهش یافته می باشند. در بین پروتئین هایی که مستقیما به جایگاه های فسفوتیروزین RTK فعال شده متصل می شوند، مولکول های رابطی وجود دارند که پروتئین های فعال کننده Ras را به کار گرفته و تحریک می کنند. زمانی که پروتئین های فعال کننده Ras تحریک می شوند، تبادل GTP با GDP را در جایگاه اتصالی نوکلئوتید گوانین در Ras تسریع می کنند. این پروتئین Ras فعال با اتصال موقتی به خانواده ای از سرین/ ترئونین کینازها، آن را تحریک می کنند که آن ها هم به نوبه خود آبشار پروتئین کیناز فعال شده با عامل میتوژن (MAP کیناز) را تحریک می کند.
زیر واحدهای آلفای گیرنده انسولین در خارج سلول قرار داشته و جایگاه اتصال انسولین هستند. اتصال لیگاند، اتوفسفریلاسیون اسید آمینه تیروزین قسمت سیتوپلاسمی زیر واحدهای بتا را القاء می کند. این اتوفسفریلاسیون اتصال پروتئین های سوبسترای سیتوزولی مثل سوبسترای 1 گیرنده انسولین (IRS-1) را تسهیل می کند. وقتی فسفریلاسیون اتفاق افتاد، این سوبسترا به عنوان پروتئین الحاقی برای پروتئین های واسطه عمل انسولین رفتار می کند. اگر چه گیرنده انسولین روی تیروزین اتوفسفریله می شود و تیروزین IRS-1 را فسفریله می کند، ولی واسطه های دیگر غالبا بر روی سرین یا ترئونین فسفریله می شوند. زیر واحد بتای فعال شده، فسفریلاسیون پروتئین های سلولی مانند اعضای خانواده IRS، Shc و Cbl را کاتالیز می کند.
گیرند های سیتوکینی:
گیرنده های مربوط به هورمون رشد، پرولاکتین، اریتروپوئیتین و سیتوکین ها فاقد فعالیت ذاتی تیروزین کینازی هستند. با اتصال سیتوکین به گیرنده، تیروزین کینازهای سیتوزولی نظیر (JAK) Janus Kinas به گیرنده متصل می شوند. اتصال JAKبه گیرنده فعال موجب فسفریلاسیون و دیمریزاسیون آن می شود. دیمرهای JAK غالبا زیر واحدهای دیگر گیرنده سیتوکینیرا در ریشه های تیروزین فسفریله می کنند. سپس این واحدهای فسفریله به عنوان جایگاه های شناسایی جهت اتصال سایر پروتئین های پیام رسان با نواحی برهمکنشSH2 (Src-Homology type-2) عمل می کنند. دیمرهای JAK به نوبه خود موجب فسفریلاسیون و دیمریزاسیون پروتئین های منومری مبدل های پیام و فعال کننده های رونویسی (STAT) آن ها می شوند. دیمرهای STAT فعال وارد هسته شده و سبب تحریک بیان برخی ژن ها می شوند.
فسفاتیدیل اینوزیتول 3-کینازها (-PI3کینازها) از طریق برهمکنش های ناحیه SH-2 با گیرنده های فاکتورهای رشد فسفریله کننده تیروزین و یا گیرنده های سیتوکینی فعال، فعال می شوند. -PI3کینازها غالبا فسفاتیدیل اینوزیتول 4، 5- بیس فسفات را فسفریله کرده و فسفاتیدیل اینوزیتول 3، 4، 5-تری فسفات (PIP3) را ایجاد می کنند. خانواده Akt / پروتئین کیناز B (PKB) به عنوان واسطه های اصلی عمل فسفاتیدیل اینوزیتول تری فسفات شناخته شده اند. پروتئین کیناز B فعال با مهار فعالیت مسیرهای پیام رسان مرگ، بقاء سلول را افزایش داده و سایر پروتئین ها و کینازهای دخیل در انتقال گلوکز و متابولیسم گلیکوژن را تنظیم می کند.
پروتئین های کوچکی به نام SOCS موجب خاتمه پیام رسانی گیرنده های سیتوکینی می شوند. این تنظیم کننده های منفی از دو مسیر عمل می کنند. اول این که، دومن SH2 بر روی پروتئین های SOCS به فسفوتیروزین های گیرنده فعال شده متصل می شود و مانع از اتصال پروتئین های پیام رسان دیگر دارای دومن SH2 (مانند STAT ها) به آن می شوند و لذا پیام رسانی گیرنده را مهار می کند. دوم این که، تمامی پروتئین های SOCS حاوی دومنی تحت عنوان جعبه SOCS هستند که اجزاء لیگاز یوبی کوئیتین E3 را فرا می خواند. برای مثال بر اثر اتصال1-SOCS، JAK2، پلی یوبی کوئیتینه شده و در پروتئازوم تجزیه می شود.
گیرنده های سرین / ترئونین کینازی:
فاکتورهای چون بتا- TGFو پروتئین های شکل دهنده استخوان (BMP) گیرنده های با فعالیت سرین / ترئونین کینازی دارند. این گیرنده ها به هنگام اتصال آگونیست های آن ها به نواحی خارج سلولی شان کمپلکس های همودیمری تشکیل می دهند. تغیرات حاصل در آرایش فضایی موجب می شود که هر زیر واحد کاتالیتیک، زیر واحد همتای خود را در ریشه سرین / ترئونین فسفریله کند. این فسفریلاسیون اولیه به هر زیر واحد همتا اجازه می دهد که پروتئین های سیتوپلاسمی به نام SMAD ها که یک خانواده چند عضوی از فاکتورهای تنظیمی ژن هستند، فسفریله و فعال کند. سپس SMAD های فعال موجب ورود کمپلکس های سیتوزولی به درون هسته و تحریک نسخه برداری برخی از ژن ها می شوند. ترومبوسپوندین (یک نوع پروتئین ماتریکس سلولی) و ماتریکس متالوپروتئازها باعث فعال شدن بتا- TGFمی شوند.
گیرنده های متصل بهG پروتئین ها (GCPR):
پروتئین های G از لحاظ عملکردی به گیرنده های هفت بار گذرنده از غشاء متصل می شوند، و به صورت کمپلکس های هترو تریمری شامل زیر واحدهای آلفا، بتا و گاما وجود دارند. زیر واحد آلفا دارای جایگاه اتصال بهGTP و فعالیت GTPase است.
پروتئین های خانوادهGs :
آدنیلات سیکلاز را تحریک کرده و موجب افزایش cAMP سلولی می شوند. cAMP بیشتر اثرات خود را به عنوان پیامبر ثانویه، با فعال کردن پروتئین کیناز وابسته به cAMP (PKA) اعمال می نماید. در غیاب cAMP، PKA یک مولکول تترامر شامل دو زیر واحد تنظیمی (زیر واحدهای R) حاوی جایگاه های اتصال به cAMP و دو زیر واحد کاتالیتیک (زیر واحدهای C) دارای فعالیت کینازی می باشد. به محض اتصال چندین مولکولcAMP به زیر واحدهای تنظیمی، تترامر تفکیک شده و به زیر واحدهای کاتالیتیک، آزاد و فعال می شود.
PKA پروتئینی به نام پروتئین متصل شونده به عنصر پاسخ به cAMP (CREB) را در ریشه سرین فسفریله می کند، این فاکتور رونویسی به هنگام فسفریله شدن به عنصر پاسخ به cAMP (CRE) که یک توالی در ناحیه تنظیمی ژن می باشد متصل می شود. بسیاری از سوبستراهای PKA، خودشان کینازهایی برای آنزیم های هدف ویژه هستند.
پروتئین های خانوادهGi :
آدنیلات سیکلاز را مهار کرده و موجب کاهشcAMP سلولی می شوند. تجزیه cAMP به وسیله نوکلئوتید فسفودی استرازها (PDEs) کاتالیز می شود که پیوند دی استری حلقوی را هیدرولیز کرده و AMP تولید می کنند. مشتقات گزانتین از قبیل تئوفیلین و کافئین، با مهارPDEs موجب افزایش میزانcAMP سلولی در غیاب تحریک هورمونی می شود.
فسفودی استراز (PDE) در سلول های استوانه ای بینایی، یک پروتئین هتروتریمریک متشکل از زیر واحدهای کاتالیتیکی آلفا و بتا و دو زیر واحد تنظیمی گاما می باشد. زیر واحد آلفای ترانسدیوسین متصل به GTP با زیر واحدهای گامایPDE کمپلکس داده و باعث جدا شدن دیمرهای آلفا و بتا کاتالیتیک فعال از PDE می شود.
توکسین ویبریوکلرا (کلراژن) با ADP -ریبوزیلاسیون زیر واحد آلفا در Gs موجب افزایش غلظتcAMP در سلول های اپی تلیال روده شده و از این طریق سبب دفع الکترولیت ها و آب از این سلول ها و ایجاد اسهال در بیماری وبا می شود.
توکسین بوردتلا پرتوسیس با ADP-ریبوزیلاسیون زیر واحد آلفا در Gi موجب افزایش غلظتcAMP سلولی و ایجاد ضایعه در بیماری سیاه سرفه می شود.
فسفولیپاز C (PLC) توسط Gq a11 فعال شده و با هیدرولیز فسفاتیدیل اینوزیتول 4، 5 بیس فسفات (PIP2) به اینوزیتول تری فسفات (IP3) و دی آسیل گلیسرول (DAG) به عنوان مولکول های پیام رسان می شوند. DAG پروتئین کینازC (PKC) را فعال کرده وIP3 هم با اتصال به گیرنده IP3در غشای شبکه آندوپلاسمی که یک کانال کلسیمی دریچه دار وابسته به IP3 است موجب آزادسازی Ca++ از شبکه آندوپلاسمی می شود و Ca++ نیز به نوبه خود سبب فعال شدن PKC می شود. مکانسیم درمانی لیتیم (Li) در درمان بیماری های روانی از طریق تداخل در مسیر فسفواینوزیتید می باشد.
گیرنده های استیل کولین (موسکارینیک)، گاسترین، کوله سیستوکینین، وازوپرسین، TRH، GnRH، آنژیوتانسین 2 و ماده P از طریق فعال کردن فسفولیپاز C، اثر خود را اعمال می کنند

انتقال پیام به واسطه فعال شدن کالمودولین:
کالمودولین (CaM) پروتئینی است در همه بافت ها بیان شده و اعمال تنظیمی Ca++ را در دامنه وسیعی از اعمال سلولی میانجیگری می کند. کالمودولین در سلول ها به شکل آزاد وجود داشته و به صورت محکم به کمپلکس های آنزیمی نظیر پروتئین کینازهای وابسته کالمودولین (CaM کینازها) متصل می شود. کالمودولین دارای چهار جایگاه اتصال به یون کلسیم (Ca++) می باشد. به محض اتصال Ca++ به کالمودولین، این پروتئین متحمل تغییراتی در کنفورماسیون فضایی خود می شود. این عمل به نوبه خود فعالیت پروتئین های مربوطه را تحت تاثیر قرار می دهد.
انتقال پیام از طریق cGMP:
فاکتور ناتریورتیک دهلیزی (ANF) به عنوان یک آتریوپپتین از طریق تحریک گوانیلات سیکلاز غشایی در سلول های هدف موجب افزایش cGMP سلولی می شود. یک شکل محلول از گوانیلات سیکلاز در بیشتر انواع سلول ها وجود دارد که دارای ساختمان هترودیمریک بوده و یک گروه پروستیک هم به آن متصل می باشد. نیتریک اکسید (NO) با اتصال به گروه هم گوانیلات سیکلاز محلول، فعالیت کاتالیتیک آن را افزایش می دهد.
در بسیاری از بافت ها به مقدار زیاد بیان می شود، اعمال می کند. تحریک PKG در عضله صاف و سلول های غیر عضلانی موجب فسفریلاسیون هماهنگ چندین پروتئین می شود که در مهار انقباض یا حرکات وابسته بهCa++ عمل می کنند. برای مثال تحریک PKG در عضله صاف موجب فسفریلاسیون و فعال شدن فسفاتازهای زنجیره سبک میوزین شده و از این طریق باعث توقف برهمکنش میوزین با اکتین می شود.
داروی سیلدنافیل سیترات با نام تجاری ویاگرا که برای درمان اختلالات نعوظی استفاده می شود با مهار انتخابی فسفودی استرازها نوکلئوتید حلقوی نوع 5 (PDE5) که در عضله صاف عروقی به میزان زیاد بیان می شود اثر خود را اعمال می نماید.
گیرنده های داخل سلولی:
گیرنده های هورمون های استروئیدی و گیرنده های غیر استروئیدی (مانند هورمون تیروئید، رتینوئیک اسید و ویتامین D3 در داخل سلول قرار دارند. گیرنده های گلوکوکورتیکوئیدی و احتمالا گیرنده آلدوسترون در سیتوپلاسم قرار دارند، در حالی که گیرنده های دیگر درون هسته و احتمالا در اتصال با کروماتین هستند.
گیرنده های هورمون های تیروئیدی (T3R) و گیرنده های رتینوئیک اسید (RAR) در حالت فاقد لیگاند به عناصر کمک سرکوبگر متصل شده و نسخه برداری ژن را مهار می کند. بعد از اتصال لیگاند، فعالیت کمک سرکوبگر این گیرنده ها کاهش یافته و به صورت کمک فعالگر عمل می کنند.
فوق خانواده گیرنده های هسته ای دارای سه دومن عملکردی اصلی شامل دومن متصل شونده به لیگاند (LBD) در ناحیه C-ترمینال، دومن متصل شونده به DNA (DBD) در قسمت میانی گیرنده که دارای دو موتیف انگشت است؛ و یک دومن آنتی ژنیک در انتهای N-ترمینال که در بین گیرنده های استروئیدی بسیار متغیر بوده و دارای یک ناحیه آنتی ژنیک مهم و یک ناحیه تعدیل کننده فعالیت نسخه برداری است. این ساختار در تمام اعضای خانواده گیرنده استروئیدی وجود دارد. گیرنده گلوکوکورتیکوئیدی به صورت یک کمپلکس است که دارای پروتئین های جدا شونده دیگری چون دیمر kDa 90 پروتئین شوک حرارتی که دومن متصل شونده به DNA گیرنده را می پوشاند و پروتئین شوک حرارتی دیگر تحت عنوان Hsp56، که یک ایمنوفیلین است و به تعدادی از داروهای قوی سرکوب کننده قوی ایمنی متصل می شود، می باشد. با اتصال هورمون به گیرنده، پروتئین های شوک حرارتی آزاد شده و کمپلکس گیرنده-هورمون به هسته انتقال می یابد.

لطفا اندیشه خود را در قسمت نظرات بنویسید.


عالی
ممنون بابت مطالب زیباتون