





















همانند سازی DNA
همتاسازی به فرایند رونوشتبرداری از یک مولکول DNA گویند.
در همانندسازی DNA، دو مولکول DNA تولید میشود که هر یک، دارای یک رشتهٔ جدید و یک رشتهٔ قدیمی هستند (ردیف نوکلئوتیدها در هر یک از مولکولهای DNA حاصل، یکسان است) که باعث میشود DNA های دختر دقیقاً مشابه دیانای مادر باشند. به این روش تکثیر که هر DNA دختر از یک رشته از DNA مادر و یک رشتهٔ نو ساخته شده است طریقهٔ نیمهحفظ شده میگویند.
چگونگی همانندسازی
دیانای پلیمراز در حال حرکت بر روی یک رشته دنا
دو رشتهٔ پلینوکلئوتیدی DNA به کمک آنزیم هلیکاز مانند زیپ از یکدیگر جدا میشوند و سپس از روی هر رشته، رشتهٔ جدیدی ساخته میشود؛ به این ترتیب که آنزیمی به نام دیانای پلیمراز بر روی نوار پلی نوکلئوتیدی حرکت میکند و با استفاده از نوکلئوتیدهای آزاد که در سیتوپلاسم وجود دارند(دارای ۲ یا ۳ گروه فسفات هستند) هر نوکلئوتید را در مقابل نوکلئوتید مکمل خود قرار میدهد.

DNAپلیمراز فقط در جهت انتهای بدون فسفات به سمت انتهای فسفات دار نوار پلی نوکلئوتیدی حرکت میکند و از انجایی که دو رشته DNA خلاف جهت هم هستند، در یک نوار همانند سازی پیوسته انجام میشود و در رشته دیگر ('۵ به '۳) به صورت قطعه قطعه انجام مشود که در نهایت آنزیم لیگاز قطعات مجاور را به هم متصل می کند (پیوند فسفو دی استر تشکیل می دهد) برای درک راحت تر این مطلب میتوانید از انیمیشن های سایت آپارات،رشد و ... استفاده کنید.
آنزیم دیانای پلیمراز توانایی دیگری نیز دارد و آن ویرایش است: در صورتی که نوکلئوتید اشتباهی به دیانایهای دختر اضافهشود، یعنی مکمل نباشد، این آنزیم برمیگردد و نوکلئتید غلط را جدا و آن را با نکلئوتید صحیح تعویض میکند. اگر این اشتباهات تصحیح نشوند در دیانایهای دختر باقی میماند و به نسل بعد منتقل میشود. به این اشتباهات تصحیح نشده جهش میگویند.
دوراهی همانندسازی
در باکتریها که دیانای حلقوی دارند همانندسازی معمولا از دو دوراهی همانندسازی شروع میشود. این دوراهیها در یک نقطهٔ خاص به نام جایگاه اغاز همانند سازی بهوجود میآیند. دوراهیهای همانندسازی به تدریج از هم دور میشوند، تا این که در نقطهٔ مقابل جایگاه اغاز همانند سازی به هم می رسنند.
دوراهی همانندسازی
در سلولهای یوکاریوتی (دوهسته ای)، هر کروموزوم از یک مولکول دیانای طویل تشکیل شده است. اما طول دنا آنقدر زیاد است که اگر قرار باشد یک کروموزوم انسان، مانند باکتری همتاسازی را از یک نقطه آغاز کند، همتا هر کروموزوم ۳۳ روز طول میکشید! از این رو همتا در سلولهای یوکاریوتی و انسان در نقاط مختلف انجام میشود(در چند نقطه). دوراهیهای همانندسازی مختلف، سبب میشوند تا یک کروموزوم انسان در حدود ۸ ساعت به طور کامل همتاسازی کند.
پیشرفتهایی که در سده اخیر نصیب علم ژنتیک شده است، تا حدود زیادی مرهون مطالعه و بررسی وراثت در باکتریها است. امروزه ثابت شده است که مکانیسمها ژنتیکی در باکتریها از نظر واکنشهای شیمیایی مشابه یاختههای یوکاریوت است. پروکاریوتها موجودات ساده و مناسبی برای بررسیهای ژنتیکی هستند. زیرا در آنها تنها یک مولکول DNA در هر یاخته وجود دارد و این DNA دارای ساختار کروموزمی پیچیدهای نیست. استفاده از میکروارگانیسمها به عنوان ابزار مطالعه ژنتیکی دارای نقاط ضعفی نیز است.
اول آنکه کوچکی اندازه این موجودات بررسی ویژگیهای ظاهری هر یاخته را دشوار میسازد. دوم آنکه تولید مثل جنسی در این موجودات وجود ندارد و یا بطور ناقص دیده میشود. پس از اینکه ساختار مولکولی DNA که نخستین بار بوسیله واتسون و کریک معرفی و ارائه شد، نحوه بیوسنتز آن را نیز در یاخته مشخص کردند. در اواخر سالهای 1950 ، کریک اصل بنیادی را مطرح کرد. این اصل بیان کننده چگونگی انتقال اطلاعات ژنتیکی از مولکول DNA به RNA و ترجمه آن در پروتئینها است.
همانندسازی DNA
در مطالعات اولیه برای همانندسازی سه الگو مطرح شد که شامل الگوهای حفاظتی ، نیمه حفاظتی و پراکنده است. در الگوی حفاظتی از روی مارپیچ دو رشتهای DNA ، یک مولکول کامل DNA ساخته میشود. در الگوی نیمه حفاظتی ابتدا دو رشته DNA از هم باز شده و در مقابل هر یک از رشتهها ، رشته مکمل ساخته میشود. در الگوی پراکنده ابتدا مولکول DNA به قطعاتی تقسیم میگردد و هر یک از قطعه رشته مکمل خود را سنتز میکند. واتسون و کریک با پژوهشهای خود بر روی مولکول DNA ، الگوی نیمه حفاظتی را منطقی و تنها راه همانند سازی میدانستند. سپس مزلسون و استال با انجام آزمایشهای بسیار ظریف و مهم ، درستی چنین الگویی را به اثبات رساندند.
آزمایش مزلسون و استال
مزلسون و استال برای اثبات فرآیند همانند سازی آزمایشی انجام دادند که به شرح زیر میباشد. آنها ابتدا یاختههای باکتری اشرشیاکلی را در محیط کشت ویژهای که نیتروژن آن از نوع سنگین (N15) بود، برای زمان معین کشت دادند و سپس یاختهها را به محیط کشت عادی که نیتروژن آن از نوع سبک (N14) بود، انتقال دادند و در محدودههای زمانی معین از یاختههای نسلهای اول ، دوم و سوم حاصل از محیط کشت جدید ، نمونه برداری کرده و DNA آنها را به روشهای اختصاصی جدا ساختند. نمونههای DNA بر روی گرادیان (شیب) چگالی کلرور منیزیم سانتریفوژ شده و در این روش ترکیبات مختلف بر اساس چگالی آنها جدا سازی میشوند.
بدین ترتیب DNA واجد وزنهای متفاوت از یکدیگر جدا میشوند. DNA معمولی که N14 دارد (DNA سبک) به علت داشتن چگالی کمتر در بالای لوله قرار میگیرد. در حالی که مولکول DNA با (N15 سنگین) در محلی پایین تر از DNA سبک واقع میشود. DNA های واجد مقادیر متفاوت N15 و N14 نیز در بینابین این دو حد جای میگیرند.
با کشت یاختههای دارای DNA واجد نیتروژن سنگین در محیط کشت حاوی نیتروژن سبک مشاهده میشود که مولکول DNA ماهیت سبک - سنگین پیدا میکند. یعنی دو رشته DNA کاملا از هم باز شده و رشتههایی در تکمیل هر یک از دو رشته قبل ساخته میشود. این رشتههای جدید همگی دارای نیتروژن سبک (محیط کشت جدید) هستند. با ادامه کشت در نسلهای دوم و سوم ملاحظه میشود که از میزان DNA سبک - سنگین کم شده و به DNA سبک افزوده میشود.
نتیجه آزمایش مزلسون و استال
مزلسون و استال با چنین مشاهداتی نتیجه گرفتند که همانند سازی در مولکول DNA به طریق نیمه حفاظتی صورت میگیرد که مستلزم باز شدن دو رشته از هم و سنتز مولکول DNA جدید در مقابل هر رشته قدیم است. این پدیده به نام همانند سازی مشهور است.
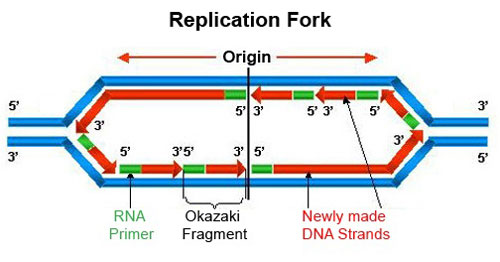
آنزیمهای لازم در همانند سازی
آنزیمهای پلیمراز
آنزیمهایی هستند که پلیمر شدن زنجیرههای پلینوکلئوتیدی را کاتالیز میکنند. تا کنون سه نوع آنزیم پلیمراز به نامهای Ι و ΙΙ و ΙΙΙ جداسازی و مشخصات آنها ارائه شدهاند. از بین آنها آنزیم پلیمراز ΙΙΙ نقش اصلی را در سنتز DNA دارد. از خصوصیات مهم آن ، این است که منحصرا نوکلئوتیدها را در جهت '5 به '3 بهم متصل میکنند و در جهت عکس نمیتواند عمل کند. آنزیم پلیمراز ΙΙ نیز در مرحلهای از سنتز DNA وارد شده و سنتز را در جهت '3 به '5 پیش میبرد. و آنزیم پلیمراز I عمل ترمیم همانند سازی را انجام میدهد.
آنزیم هلیکاز
این آنزیم به مولکول DNA دو رشتهای متصل شده و با عمل خود موجب باز شدن دو رشته از یکدیگر میشود.
آنزیم لیگاز
در مرحلهای از سنتز DNA وارد عمل شده و دو رشته DNA را بهم پیوند میدهد.
آنزیم پریماز
آنزیمی است که در ساختن قطعه کوچک RNA پرایمر ، هنگام همانند سازی وارد عمل شده و نوکلئوتیدهایی از نوع اسید ریبونوکلئوتید را به یکدیگر متصل میکند. تعدادی پروتئینهای ویژه وجود دارند که پس از باز شدن دو رشته DNA از یکدیگر به محلهای باز شده متصل شده و مانع اتصال مجدد دو رشته به یکدیگر میشوند.
همانند سازی متوالی
در روی مولکول DNA نقاطی وجود دارند که همانند سازی از آنها آغاز میشود. این نقاط مبدا همانند سازی خوانده میشوند. در DNA باکتریها ، یک مبدا همانند سازی و در DNA موجودات عالی ، تعدادی زیادی از این مبدا وجود دارند. هنگام همانند سازی ابتدا آنزیم هلیکاز به مارپیچ دو رشتهای DNA متصل شده و پیچش DNA را در آن نقطه باز میکند. پرتئینهای DBP به ناحیه باز شده هجوم آورده و با اتصال به DNA تک رشتهای مانع از جفت شدن بعدی DNA میشوند.
ناحیهای را که هلیکاز به آن متصل میشود، چنگال همانند سازی مینامند. همانند سازی به صورت دو سویه است. آنزیم پلیمراز ΙΙΙ که اتصال نوکلئوتیدها را به یکدیگر به عهده دارد، فقط میتواند همانند سازی را در جهت 3 به 5 پیش ببرد. در این حالت دو رشته مولکول DNA در خلاف جهت یکدیگر هستند. در نتیحه رشتهای که در جهت '5 به '3 سنتز میشود، به راحتی سنتز DNA را آغاز کرده و پیش میبرد. این رشته به نام رشته راهنما معروف است. در همانند سازی این رشته را متوالی مینامند.
همانند سازی نامتوالی
در مولکول DNA رشتهای که '5 آزاد دارد، سنتز DNA طبق آنچه درباره رشته راهنما ذکر شد، انجام نمیگیرد. دلیل آن این است که آنزیم پلیمراز ΙΙΙ نمیتواند نوکلئوتیدها را در جهت 3 به 5 کاتالیز کند. لذا میبایست مکانیسم دیگری برای سنتز این رشته از DNA وجود داشته باشد. این رشته DNA به نام رشته عمل کننده یا پیرو معروف است. در این حالت ابتدا دو رشته DNA در فواصل معینی از یکدیگر باز شده و آنزیم پریماز در آن محل قرار میگیرد و با استفاده از ریبونوکلئوتیدها ، RNA کوچکی ساخته میشود که RNA پرایمر نام دارد.
انتهای 3 این RNA کوچک که از روی الگوی DNA ساخته شده است، میتواند به آنزیم پلیمراز III امکان دهد تا دزاکسی ریبونوکلئوتیدها را به انتهای آن متصل کند. لذا در این رشته از مولکول DNA قطعاتی از DNA سنتز میشوند که قطعات اوکازاکی نام دارد. (اوکازاکی نخستین کسی بود که این قطعات سنتز شده DNA را با میکروسکوپ الکترونی مشاهده کرد).
در این حالت آنزیم پلیمراز I وارد عمل شده و به ترتیب یکی یکی ریبونوکلئوتیدها را در جهت 5 به 3 برداشته و به جای آنها نوکلئوتیدهای از انواع دزاکسی جایگزین میکند تا این که قطعات همه از نوع دزاوکسی شوند. سپس انتهای قطعات ساخته شده بوسیله آنزیم لیگاز به هم متصل شده و یک رشته ممتد DNA حاصل میشود. اندازه هر قطعه اوکازاکی حدود 1000 تا 2000 نوکلئوتید است.
طی همانندسازی DNA، ماده ژنتیکی سلول تکثیر می شود. آنزیم های متفاوتی در این فرایند باعث سنتز (ترکیب دو ماده اولیه و تولید یک چیز جدید) رشته های جدید از روی رشته الگو می گردد.
DNA ماده ژنتیکی هر سلول است. در تمامی مراحلی که یک سلول به دو سلول دختر تقسیم می شود و تمام اندام ها نیز می توانند کپی شوند شاهد تقسیم DNA در هسته هستیم. فرایند دو برابر شدن DNA، همانندسازی (replication) نام دارد. همانندسازی در چندین مرحله که شامل آنزیم های همانندسازی و RNA است دنبال می گردد.
در سلول های یوکاریوتی، مانند سلول های گیاهی و جانوری، همانندسازی DNA در فاز S اینترفاز چرخه سلولی انجام می شود. (اینترفاز مرحلهای از روند تقسیم است که در آن یاخته تقسیم نمیشود. در این مرحله یاخته خود را برای ورود به مرحله دیگر تقسیم آماده می کند. اینترفاز شامل سه مرحله G1 ، S و G2 است.)
ساختار DNA
دئوکسی ریبونوکلئیک اسید شامل یک قند 5 کربنه دئوکسی ریبوز، یک فسفات و بازهای آلی است.DNA شامل دو زنجیره نوکلئیک اسیدی به شکل مارپیچی است. این شکل مارپیچی به فشردگی آن کمک می کند.DNA در ساختارهای فنرمانندی به نام کروماتین قرار دارد. در طول تقسیم سلولی کروماتین به شکل کروموزوم فشرده می گردد.
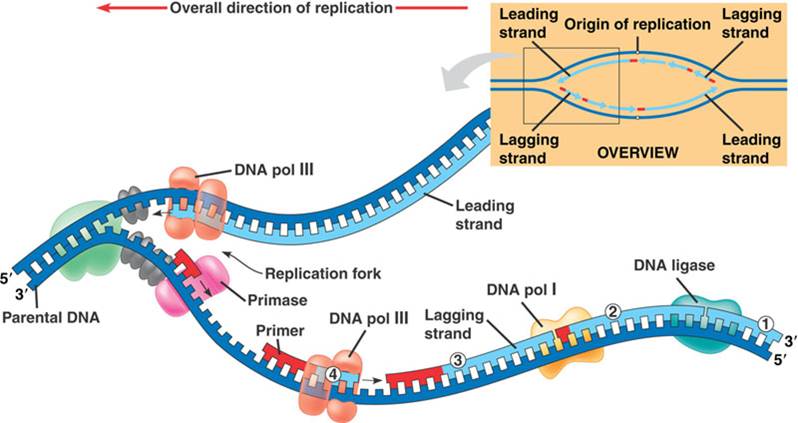
مرحله 1: تشکیل چنگال همانندسازی
قبل از این که همانندسازی شروع شود، دو رشته باید به صورت رشته های منفرد از هم باز شوند. DNA از 4 باز آلی تشکیل شده است: آدنین (A)، تیمین (T)، سیتوزین (C) و گوانین (G).
آدنین تنها با تیمین جفت می شود وگوانین با سیتوزین. برای آغاز همانندسازی اینها باید باز شوند. این عمل به وسیله آنزیم DNA هلیکاز صورت می گیرد. این آنزیم پیوندهای هیدروژنی بین بازها را برای جدا کردن رشته ها و ایجاد یک شکل Y مانند به نام چنگال همانندسازی می شکند. این منطقه الگوی شروع همانندسازی است.
DNA در هر دو رشته دوطرفه است: انتهای 5' و انتهای 3'. سمت 5' در اسکلت اصلی دارای گروه فسفات و انتهای 3' دارای گروه هیدروکسیل (OH) است. این دوطرفه بودن برای همانندسازی مهم است و باعث پیشرفت همانندسازی از سمت 5 به 3 می گردد. اگرچه چنگال همانندسازی دوطرفه است ولی یک رشته در جهت 3 به 5 سازمان دهی شده است و رشته پیشرو (leading strand) و رشته دیگر در جهت 5 به3 سازمان دهی شده است و رشته پیرو (lagging strand) نام دارد. دو رشته در دو مسیر متفاوت همانندسازی می شوند.
مرحله 2: اتصال پرایمرها
قطعه کوتاهی از RNA به نام پرایمر به انتهای 3 رشته متصل می گردد. پرایمر همیشه به نقطه شروع همانندسازی متصل می شود.
مرحله 3: طویل سازی
آنزیم DNA پلیمر مسئول ایجاد رشته های جدید در طی طویل سازی است. 5 نوع متفاوت DNA پلیمر در باکتری ها و انسان وجود دارد. در باکتری ای کولای (E. coli)، پلیمراز III آنزیم اصلی همانندسازی است در حالی که پلیمراز I، II، IV و Vمسئول تعمیر و تصحیح خطا است. بنابراین پلیمراز III به رشته ها در محل پرایمر متصل می شود و شروع به اضافه کردن بازهای جدید می کند. چون همانندسازی درجهت 5 به 3 انجام می شود، رشته های جدید پیوسته هستند.
در سلول های یوکاریوتی پلیمراز آلفا، دلتا و اپسیلون باعث شروع همانندسازی می گردند. رشته پیرو به وسیله اتصال به پرایمرهای چندگانه همانندسازی می کند. پرایمر شامل چند باز است. DNA پلیمراز، قطعاتی از DNA را به نام قطعات اوکازاکی به رشته ها اضافه می کند. این بخش از فرایند همانندسازی غیرپیوسته است و در پایان این قطعات به هم متصل می شوند.
مرحله 4: پایان
پس از تشکیل رشته های پیوسته و غیر پیوسته، آنزیم اگزونوکلئاز قطعات پرایمر را از ابتدای رشته ها حذف می کند. اگزونوکلئازهای دیگر وظیفه بررسی و حذف اشتباه ها را دارند. آنزیم دیگری به نام DNA لیگاز قطعات اوکازاکی را به هم متصل می کند. انتهای رشته های والدی، شامل قطعاتی از توالی DNA است و تلومراز نام دارد. تلومراز یک عامل حفاظتی در انتهای کروموزوم است و از الحاق کروموزوم ها جلوگیری می کند. آنزیم تلومراز باعث سنتز تلومر در انتهای DNA می گردد. در پایان همانندسازی دو مولکول DNA تولید می کند که دارای یک رشته از مولکول پدری و یک رشته جدید است.
آنزیم های مسئول همانندسازی در سلول های یوکاریوتی، آنزیم هایی مسئول همانندسازی هستند که عبارتند از:
DNA هلیکاز: جداکردن دو رشته DNA و حرکت در طول DNA
پرایماز: RNA پلیمرازی که قطعات پرایمرهای RNA را تولید می کند. پرایمرها به عنوان نقطه شروع همانندسازی عمل می کنند.
DNA پلیمراز: سنتز مولکول های DNA جدید با اضافه کردن نوکلئوتیدها به رشته های رهبر و پیرو
توپوایزومراز: آنزیم شکست و بست، از اتصال مجدد رشته ها و تشکیل سوپرکویل جلوگیری می کند.
اگزونوکلئازها: آنزیم هایی که بازهای نوکلئوتیدی را از انتهای زنجیره DNA حذف می کند.
DNA لیگاز: قطعات DNA را با تشکیل باندهای فسفودی استری به هم متصل می کنند.
مدتها قبل از آنکه ساختمان DNA شناخته شود دانشمندان از توانایی موجود زنده در ایجاد نسخه های دقیقا مشابه خود در تعجب بودند.
کشف قوانین مندل در سال 1900 میلادی سبب تولید علم ژنتیک گردید و طی سالهای 1952 تا 1962 ساختمان DNA درخشید، کد ژنتیکی شکوفا گردید و فرایندهای رونویسی و ترجمه شرح داده شدند.
در سالهای 1971 تا 1973 یک متدولوژی کاملا جدید توسعه یافت و امکان داد تا آزمایشهایی را که قبلا عملی نبودند با موفقیت طرح ریزی و انجام شوند. این روش تحت عنوان علم بیو تکنولوژی نامیده شد.
اما در این علم توجه اذهان عمومی بیشتر به وسیله مشابه سازی بخصوص مشابه سازی انسان جلب شد. با پیشرفت این علم و نتایج حاصل از تحقیقات دانشمندان نشان می داد که از یک سلول در حال تقسیم جنین وزغ یا قورباغه می توان دسته ای از این موجود را که دارای ساختار ژنتیکی یکسانی باشند، خلق کرد.
مفهوم همانند سازی
کلون عبارتست از گروهی از موجودات زنده اعم از ژن، سلول، ویروس و... که از نظر ژنتیکی مشابه هستند.
همانند سازی یا کلونینگ به سه روند مجزا از یکدیگر اطلاق می شود که در ذیل به بررسی آنها می پردازیم:
متداولترین نوع آن شکل جنینی آن است، بدین صورت که یک سلول تخم به دو سلول تقسیم میشود و هرکدام از این در سلول درصورت قرار گرفتن در رحم مادر، انسان مشابه دیگری ایجاد خواهند کرد.
نوع دیگر همانند سازی استفاده از سلول بالغ است. بدین صورت که هسته یک سلول تخمک را خارج می کنیم و هسته یک سلول بالغ را جایگزین آن می کنیم، در نتیجه ژن های خاموش فعال می شوند و انسانی با مشخصات ژنتیکی فرد بالغ تولید خواهد کرد.
نوع سوم همانندسازی شیوه درمانی آن است. با استفاده از این روش می توان سلول های پوست فردی را که نیاز به سلول عصبی دارد، طوری برنامه ریزی کرد که سلول عصبی بسازد.
همانند سازی DNA
برای همانند سازی سه الگو مطرح است: الگوی حفاظتی، نیمه حفاظتی و پراکنده
در الگوی حفاظتی از روی مارپیچ دو رشته ای DNA یک مولکول کامل DNA ساخته می شود.
در الگوی نیمه حفاظتی، دو رشته از هم باز شده و در مقابل هریک از رشته ها رشته مکمل ساخته می شود.
در الگوی پراکنده ابتدا مولکول DNA به قطعاتی تقسیم می شود و هریک از قطعات رشته مکمل خود را سنتز می کند.
واتسون و کریک با پژوهش های خود، الگوی نیمه حفاظتی را تنها راه همانند سازی می دانستند. مزلسون و استال نیز با انجام آزمایشاتی درستی چنین الگویی را اثبات کردند.
آزمایش مزلسون- استال
آنها یاخته های باکتری E-Coli را در محیط کشت ویژه ای که نیتروژن آن از نوع سنگین (N15) بود، کشت دادند و سپس یاخته ها را به محیط کشتی که نیتروژن آن از نوع سبک (N14) بود، انتقال دادند. سپس از یاخته های نسل اول، دوم و سوم حاصل از محیط کشت جدید نمونه برداری کرده و DNA آنها را جدا کردند و بر روی گرادیان(شیب) چگالی کلرور منیزیم سانتریفوژ کردند. بدین ترتیب DNA دارای وزن های متفاوت از یکدیگر جدا می شوند.
DNA سبک که (N14) دارد به علت داشتن چگالی کمتر در بالای لوله قرار می گیرد ولی DNA سنگین (N15) به دلیل داشتن چگالی بیشتر پایینتر از DNA سبک قرار می گیرد. DNA دارای مقادیر متفاوت N14و N15 نیز در بین این دو جای می گیرند.
با کشت یاخته های دارای DNA سنگین در محیط کشت حاوی نیتروژن سبک مشاهده می شود که مولکول DNA ماهیت سبک- سنگین پیدا می کند یعنی دو رشته DNA از هم باز شده و رشته هایی در تکمیل هریک از دو رشته قبل ساخته می شود، این رشته ها همگی دارای نیتروژن سبک هستند. با ادامه کشت در نسل های دوم و سوم ملاحظه می شود که از میزان DNA سبک- سنگین کاسته شده و به DNA سبک افزوده می شود.
نتیجه آزمایش مزلسون و استال
مزلسون و استال با چنین مشاهداتی نتیجه گرفتند که همانندسازی به طریق نیمه حفاظتی صورت می گیرد که مستلزم باز شدن دو رشته از هم و سنتز مولکول DNA
جدید در مقابل هر رشته قدیم است. این پدیده به نام همانندسازی مشهور است.
آنزیم های لازم در همانند سازی
DNA پلی مرازها
DNA پلی مرازها افزودن نوکلیوتیدها طی طویل شدن زنجیره را کاتالیز می کند. این آنزیم ها برای عمل هم به الگوها((Template وهم به پرایمرها(Primers) نیازمند بوده و از5´-DNTP استفاده می کنند.
تکنیک های قوی آزمایشگاهی مربوط به تعیین توالی DNA و واکنش زنجیره پلی مراز، هر دو بر اساس نیاز به الگو و پرایمر برای پلیمریزه شدن DNA می باشد.
سلول ها دارای DNA پلی مرازهای مختلفی هستند که اعمال ویژه ای بر عهده دارند، این اعمال شامل جنبه های مختلفی از همانند سازی و ترمیم آسیب به رشته الگو می باشد.(جدول کتاب دولین).
DNA پلی مرازهایی که در همنند سازی شرکت می کنند از دو طریق به تضمین دقت کمک می کنند:
1- با انتخاب اولیه نوکلیوتید مناسب برای افزودن
2- با تصحیح آ نزیمی
انتخاب اولیه بر اساس صحت نوکلیوتید تازه وارد به جایگاه فعال فعال پلی مراز می باشد. یک نوکلیوتید تازه وارد که با نوکلیوتید موجود در رشته الگو، پیوند هیدروژنی صحیح برقرار می کند می تواند به زنجیره در حال رشد اضافه شود.
یک نوکلیوتید تازه وارد که پیوند هیدروژنی صحیح برقرار نمی کند، برای کاتالیز شدن مناسب نیست. این امر سبب می شود میزان اشتباهات از 10-4 به 10-6 برسد.
دومین راه افزایش دقت توسط DNA پلیمرازها،عمل تصحیح ((Proofreading است. این عمل با فعالیت اگزو نوکلیازی 3´ به 5´ انجام می شود که نوکلیوتید های اشتباه جفت شده را از انتهای 3´ رشته در حال رشد برمی دارد.
تا کنون سه نوع آنزیم DNA پلی مراز به نام هایIII,II,I شناخته شده اند:
از بین آنها آنزیم DNA پلی مراز III نقش اصلی را در سنتز DNA دارد و از خصوصیات مهم آن، این است که منحصرا نوکلیوتیدها را در جهت 5´ به 3´ به هم متصل می کند و در جهت عکس نمی تواند عمل کند.
DNA پلی مراز II نیز در مرحله ای از سنتز DNA وارد شده و سنتز را در جهت 3´ به 5´ پیش می برد.
و آنزیم DNA پلی مراز I عمل ترمیم و همانند سازی را انجام می دهد.
آنزیم های هلیکاز(Helicase)
جدا شدن دو رشته والد DNA از یکدیگر این امکان را فراهم می سازد تا هر رشته به عنوان الگو برای سنتز رشته جدید مورد استفاده قرار گیرد. جدا شدن دو رشته والد DNA باعث ایجاد ساختاری موسوم به چنگال همانند سازی می گردد. این جدا شدن نیازمند صرف انرژی قابل توجهی است بطوریکه تنها در دماهای بالا و معمولا بالاتر از 90°C رخ می دهد.
سلول ها برای جداسازی رشته های والد در دماهای فیزیولوژیکی از آنزیم هایی به نام هلیکازها (Helicase) استفاده می کنند. هلیکازها به DNA تک رشته ای متصل می شوند و در طول DNA در یک جهت ثابتی حرکت می کنند.
آنزیم های هلیکاز برای عملکرد در هر مرحله نیازمند هیدرولیز ATP هستند.
هلیکاز با هل دادن DNA والد، چنگال همانند سازی را می سازد.
همانندسازی متوالی
در روی مولکول DNA نقاطی وجود دارد که همانند سازی از آنها آغاز می شود. این نقاط مبدا همانند سازی نامیده می شوند. در DNA باکتری ها یک مبدا همانند سازی و در DNA موجودات عالی، تعداد زیادی از این مبدا وجود دارند.
هنگام همانند سازی ابتدا آنزیم هلیکاز به مارپیچ دو رشته ای DNA متصل شده و پیچش DNA را درآن نقطه باز می کند. پروتیین های DBP به ناحیه باز شده هجوم آوره و با اتصال به DNA تک رشته ای مانع از جفت شدن بعدی DNA می شود. ناحیه ای را که هلیکاز به آن متصل می شود، چنگال همانند سازی می نامند.
همانند سازی به صورت دو سویه است و همانطور که گفته شد آنزیم پلی مرازIII که اتصال نوکلیوتیدها را به یکدیگر به عهده دارد فقط می تواند همانند سازی را در جهت 5´ به 3´ پیش ببرد. در این حالت دو رشته مولکول DNA در خلاف جهت یکدیگر هستند، در نتیجه رشته ای که در جهت 5´ به 3´ سنتز می شود براحتی سنتز DNA را آغاز کرده و پیش می برد.
این رشته به نام رشته راهنما معروف است. در همانند سازی این رشته را متوالی می نامند.
همانند سازی نا متوالی
در مولکول DNA ای که رشته 5´ آزاد دارد، سنتز DNA طبق آنچه درباره رشته راهنما ذکر شد، انجام می گیرد. دلیل آن این است که آنزیم پلی مراز III نمی تواند نوکلیوتیدها را در جهت 3´ به 5´ کاتالیز کند، لذا می بایست مکانیسم دیگری برای سنتز این رشته از DNA وجود داشته باشد. این رشته DNA به نام رشته عمل کننده یا پیرو معروف است. در این حالت ابتدا دو رشته DNA در فواصل معینی از یکدیگر باز شده وآنزیم پرایمز در آن حل قرار می گیرد و با استفاده از ریبونوکلیوتید ها، RNA کوچکی ساخته می شود که RNA پرایمر نام دارد. انتهای 3´ این RNA کوچک که از روی الگوی DNA ساخته شده است، می تواند به آنزیم پلی مراز III این امکان را بدهد تا دزاکسی ریبو نوکلیوتیدها را به انتهای آن متصل کند. لذا در این رشته از مولکول DNA قطعاتی از DNA سنتز می شودکه قطعات اوکازاکی نام دارد.
در این حالت آنزیم پلی مراز I وارد عمل شده و به ترتیب یکی یکی ریبونوکلیوتید ها را در جهت 5´ به 3´ برداشته و بجای آنها نوکلیوتید های از انواع دزوکسی جایگزین می کند تا اینکه همه قطعات از نوع دزوکسی شوند. سپس انتهای قطعات ساخته شده بوسیله آنزیم لیگاز به هم متصل شده و یک رشته ممتد DNA حاصل می شود. اندازه هر قطعه اوکازاکی حدود 1000 تا 2000 نوکلیوتید است.
همانند سازی DNA در یوکاریوتها
در یوکاریوتها، کروموزوم ساختار پیچیده ای است که در غشای هسته ای محصور شده است. تعداد کرموزوم ها در هر گونه ثابت است. کروموزوم ها علاوه بر DNA حاوی هیستونها، مقداری RNA و پروتیین های غیر هیستونی هستند.
طرح همانند سازی در DNA یوکاریوت ها همانند پروکاریوتهاست و فقط در جزییات تفاوت دارند.
از آنجا که کروموزوم های یوکاریوتی دارای مقدار زیادی DNA هستند، لذا رپلیکون های فراوانی دارند و بنابراین سنتز DNA می تواند از چندین محل آغاز شود. اگر چنین وضعی رخ نمی داد، مرحله S در چرخه زندگی یاخته می بایست بسیار طولانی شود. در انسان یک تک یاخته دیپلویید 46 کروموزوم دارد که طول DNA در هریک 2000 تا 3000 میکرومتر است. بدیهی است که رپلیکونهای فراوانی وجود دارند، بنابراین مرحله S تا 8 ساعت طول می کشد. اگر یک رپلیکون وجود داشت، حدود 230 روز طول می کشید تا همانند سازی در مرحلهS انجام شود. همه رپلیکون ها نمی توانند بطور همزمان DNA بسازند، از این رو همانند سازی 6 تا 8 ساعت طول می کشد.
همانند سازی از نقطه مرکزی مبدا در هر رپلیکون آغاز می شود و در دو جهت پیش می رود. نقطه آغازی بصورت دو راهی است و از الحاق این دو راهی ها تعداد زیادی حباب ایجاد می شود .
برعکس DNA عریان در پروکاریوتها، کروماتین در کروموزومهای یوکاریوتها به شکل نوکلیوزوم هاست که DNA در آنها بسته بندی شده است. پیشنهاد شده است که نوکلیوزوم همراه با DNA ای که بخشی از آن باز شده، به دو نیمه مشخص تقسیم می شود و بدین ترتیب همانند سازی آغاز می شود. پس از تکمیل همانند سازی، هر نیمه نوکلیوزوم سریعا با نوکلیوزوم نوساخته که در آن هیستونها و DNA های جدید ساخته شده اند، همراه می گردد. بنابراین یک نوکلیوزوم کامل متشکل از یک نیمه قدیمی و یک نیمه جدید ساخته می شود.
مکانیسم همانندسازی در یوکاریوتها مانند یکدیگر است. در یوکاریوتها نیز همانند سازی DNA بصورت نیمه حفاظتی است و قطعات اوکازاکی که بوسیله RNA های آغازی شروع می شوند، در این امر دخالت دارند. طول RNAی آغازی در حدود 10 نوکلیوتید است.
انتهای 3´ هیدروکسیل قطعه اوکازاکی بوسیله DNA پلی مراز دراز می شود، این قطعات در DNA یوکاریوتها کوتاهند و از 100 تا 200 نوکلیوتید تشکیل شده است. پس از تشکیل قطعات اوکازاکی، RNAی آغازی برداشته می شود و فاصله حاصل با قطعه ای از DNA بوسیله DNA پلی مراز I پر میگردد. سپس قطعات به کمک DNA لیگاز به یکدیگر متصل می شود.
عکس شماره 1 : دریافت
عکس شماره 2 : دریافت
عکس شماره 3 : دریافت
برای دریاف فایل PDF کلید کنید : دریافت
