





















تروپومیوزین
تروپومیوزین آلفا-مارپیچ پروتئین مارپیچ دو رشته ای موجود در شبکه رشته ای سلول ها است.
میوزین و اکتین اسکلت
تمام موجودات زنده شامل ساختارهایی هستند که تمامیت جسمی به سلول های خود را دارند. این سازه در مجموع به عنوان اسکلت سلولی شناخته شده است، و یکی از مهمترین و سیستم های قدیمی پلیمر رشته اکتین پروتئین است. ساختار دوم از پروتئین، تروپومیوزین ، بخشی جدایی ناپذیر از ترین رشته های اکتین در حیوانات است.
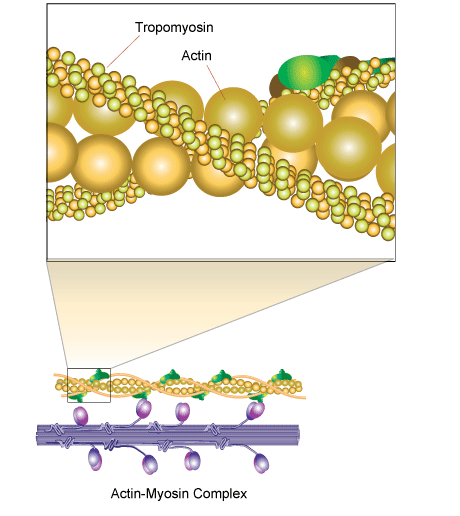
تروپومیوزین یک خانواده بزرگ از اجزای جدایی ناپذیر از رشته های اکتین است که نقش مهمی در تنظیم عملکرد رشته های اکتین در هر دو عضله و سلول های غیر ماهیچه ای بازی می کند. این پروتئین ها حالت های مختلفی دارند از میله ای شکل گرفته تا ساختارهای داغ سیم پیچ یا هومو-دیمر که در طول شیار α-مارپیچ از ترین رشته های اکتین دروغ است. تعامل در امتداد طول رشته اکتین رخ می دهد.
تروپومیوزین اغلب به دو گروه، عضله ایزوفرم تروپومیوزین و ایزوفرم تروپومیوزین سلول های غیر ماهیچه ای دسته
بندی می شوند.
ایزوفرم تروپومیوزین عضلات در تنظیم فعل و انفعالات
بین اکتین و میوزین در سارکومر عضلانی درگیر و نقش محوری در انقباض عضله تنظیم بازی
کند. ایزوفرم تروپومیوزین سلول های غیر ماهیچه ای در تمام سلول عمل، هر دو عضله و سلول های غیر ماهیچه ای سلول،
و در طیف وسیعی از مسیرهای سلولی که کنترل و تنظیم اسکلت سلولی سلول و دیگر توابع کلیدی
سلولی شرکت دارند.
سیستم اکتین رشته است که در تنظیم این مسیرهای سلولی درگیر پیچیده تر از سیستم های رشته اکتین که تنظیم عضلات است. سیستم انقباضی اکتین متکی بر 4 ایزوفرم رشته و 5 ایزوفرم تروپومیوزین ، در حالی که سیستم های اکتین رشته از اسکلت سلولی با استفاده از 2 ایزوفرم اکتین رشته و بیش از 40 ایزوفرم تروپومیوزین.
نقش های خاص و عملکرد ها
نقش در در انقباض عضلانی اسکلتی
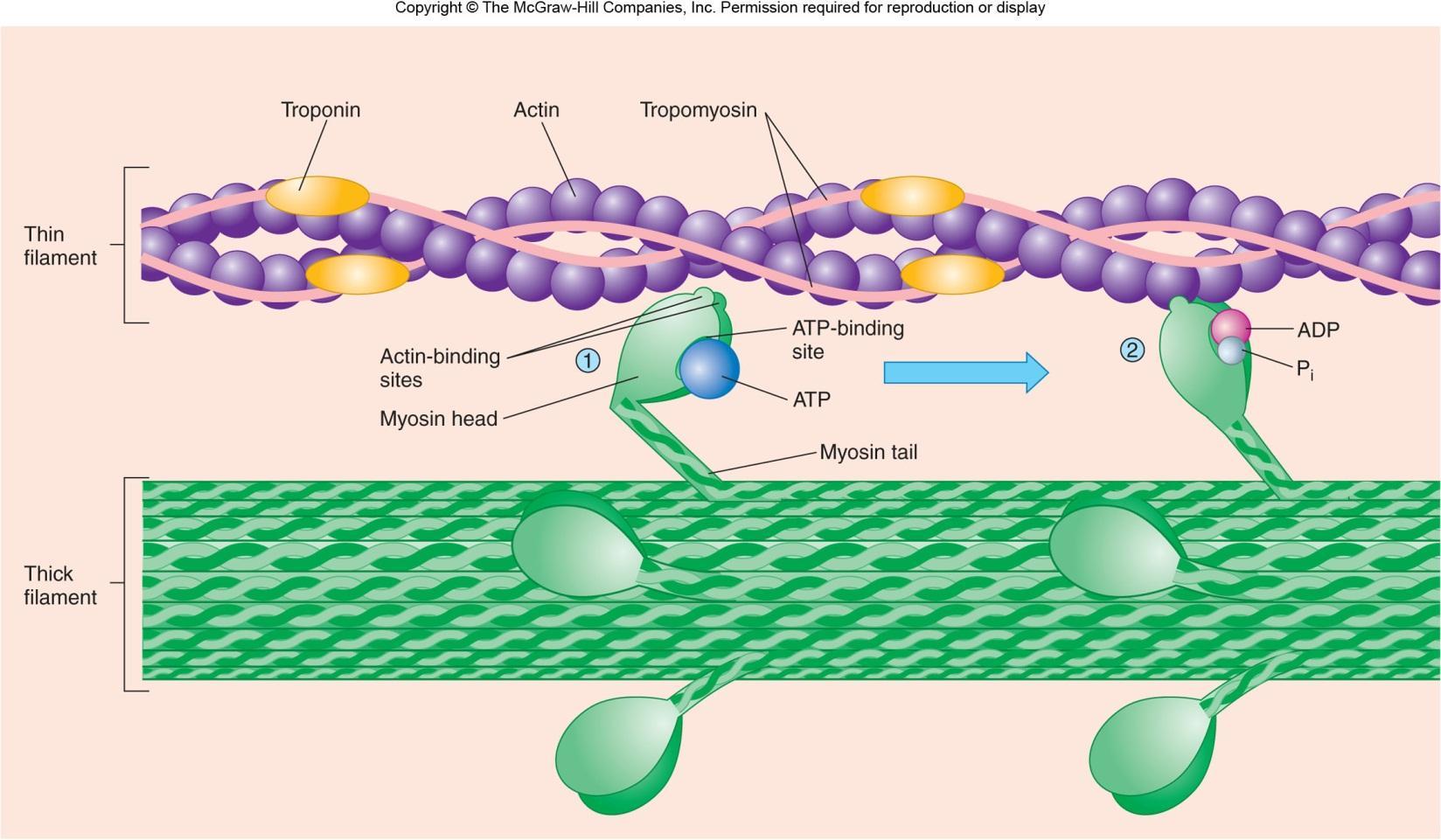
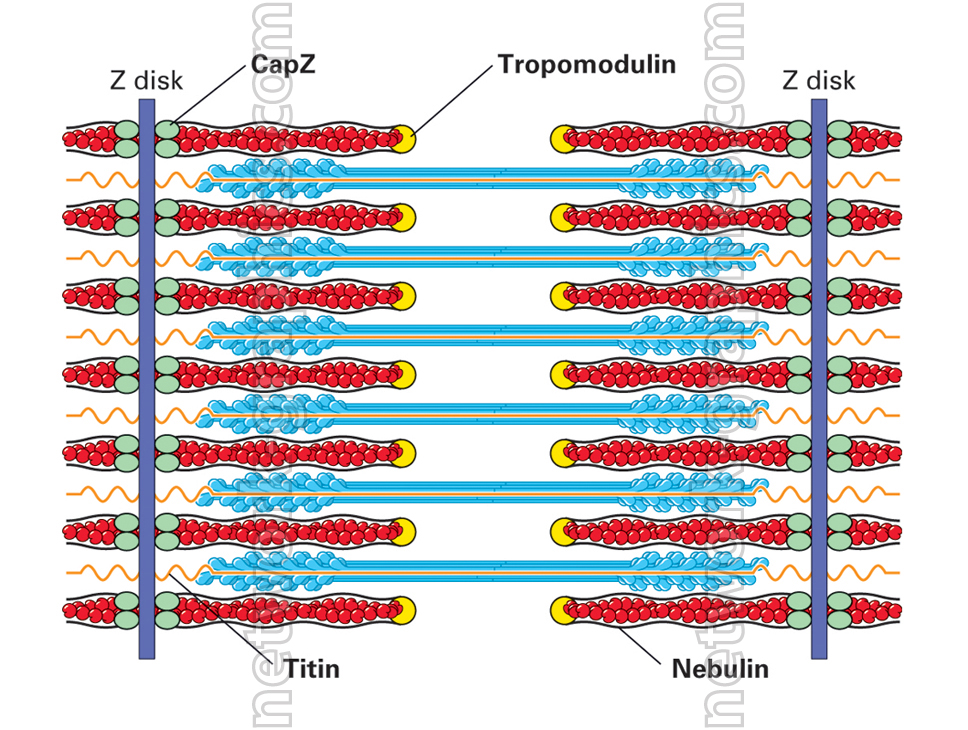
عضله است بزرگ، سلول های چند هسته (فیبرهای عضلانی) تشکیل شده است. هر فیبر عضلانی است که با آرایه های طولی از تارچه بسته بندی شده. تارچه از تکرار ساختارهای پروتئینی یا سارکومر، واحد عملکردی عضلات اسکلتی تشکیل شده است. سارکومر یک آرایه پروتئین بسیار ساختار است، متشکل از interdigitating رشته ضخیم و نازک، که در آن رشته های باریک و افسار را به یک ساختار پروتئین، Z-خط هستند. تعامل پویا میان رشته ضخیم و نازک منجر به انقباض عضله.
میوزین متعلق به یک خانواده از پروتئین موتور، و ایزوفرم های عضلانی از این خانواده شامل رشته ضخامت دارد. رشته نازک از ایزوفرم عضله اسکلتی اکتین ساخته شده است. هر پروتئین میوزین، به پایان می رسد، در امتداد رشته های اکتین نازک، بارها و بارها با اتصال به سایت های میوزین متصل شونده به همراه رشته اکتین، تشدید و رها. در واقع، حرکت رشته ضخیم و یا اسلاید در امتداد رشته نازک، و در نتیجه انقباض عضلات. این فرایند به عنوان مدل رشته کشویی شناخته شده است.
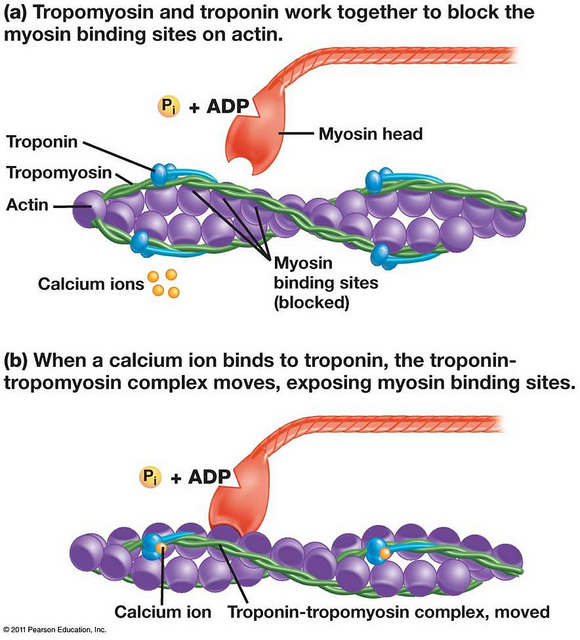
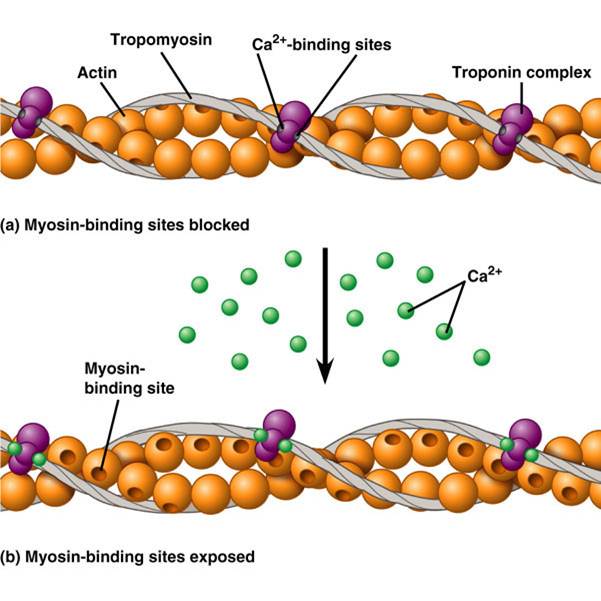
اتصال سر میوزین به اکتین عضلانی یک فرایند بسیار تنظیم است. رشته نازک اکتین، تروپومیوزین، و تروپونین ساخته شده است. انقباض عضله اسکلتی است تکانه های عصبی که به نوبه خود باعث تحریک آزادسازی های Ca2 + باعث شده است. انتشار های Ca2 + از شبکه سارکوپلاسمی باعث افزایش غلظت های Ca2 + در سیتوزول. یون های کلسیم و سپس به تروپونین، است که با تروپومیوزینهمراه متصل شوند. اتصال تغییرات علل به شکل تروپونین و پس از آن باعث ایزوفرم تروپومیوزینبه تغییر موضع خود را در رشته اکتین. این تغییر در موقعیت معرض سایت های میوزین اتصال در رشته های اکتین، اجازه می دهد سر میوزین از رشته های ضخیم برای اتصال به رشته نازک.
مطالعات ساختاری و بیوشیمیایی نشان می دهد که موقعیت تروپومیوزینو تروپونین در رشته نازک تعامل بین سران میوزین از رشته ضخیم و محل های اتصال در اکتین از رشته نازک تنظیم می کند. پراش اشعه X و میکروسکوپ خاص نشان می دهد که تروپومیوزین بلوک sterically دسترسی به میوزینرشته اکتین.

اگر چه این مدل به خوبی تثبیت، معلوم نیست که آیا حرکت تروپومیوزینبه طور مستقیم باعث سر میوزین به شرکت رشته اکتین است. به این ترتیب، یک الگوی جایگزین پدید آمده است، به موجب آن جنبش از تروپومیوزیندر توابع رشته به عنوان یک سوئیچ آلوستریک است که با فعال میوزین اتصال مدوله می کند، اما تنها با تنظیم میوزین اتصال عمل نمی کند.
تنظیم انقباض در عضله صاف
عضله صاف یک نوع از عضلات مخطط غیر است، و بر خلاف عضله مخطط، انقباض عضله صاف است تحت کنترل آگاهانه نیست. عضله صاف ممکن است خود به خود و یا منظم منقبض می شود و توسط تعدادی از عوامل فیزیکوشیمیایی (هورمون ها، مواد مخدر، انتقال دهنده های عصبی) ناشی از. عضله صاف است که در داخل دیوار اندام های مختلف و لوله در بدن مانند عروق مری، معده، روده ها، برونش، مجرای خروجی مثانه، مثانه، و خون یافت می شود.
اگر چه عضلات صاف انجام آرایه به طور منظم از رشته های ضخیم و نازک مانند سارکومر عضلات مخطط را شکل داد، انقباض است که هنوز هم به دلیل همان مکانیزم کشویی رشته های crossbridges میوزین تعامل با رشته های اکتین کنترل می شود. رشته نازک از عضله صاف از اکتین، tropomyosin، caldesmon و کالمودولین ساخته شده است. در این نوع از عضلات، caldesmon و کالمودولین کنترل انتقال tropomyosin واسطه بین روشن و خاموش فعالیت. Caldesmon متصل به اکتین، tropomyosin، کالمودولین و میوزین، که تعامل با اکتین مهم ترین است. اتصال caldesmon است به شدت توسط tropomyosin تحت تاثیر قرار. Caldesmon یک مهار کننده actinomyosin ATPase و تحرک است، و هر دو اکتین اتصال و مهار caldesmon تا حد زیادی در حضور tropomyosin افزایش یافته است.
انقباض عضله صاف است و با آزادی های Ca2 + آغاز می شود. CA2 + متصل به و فعال کالمودولین، که پس از آن متصل به caldesmon. این اتصال باعث پروتئین caldesmon به خلاص از رشته اکتین، میوزین افشای سایت های الزام آور در رشته اکتین. میوزین سر موتور توسط میوزین کیناز نور زنجیره ای، اجازه می دهد سر میوزین برای تعامل با رشته های اکتین و علت انقباض فسفوریله.
نقش در عملکرد اسکلت سلولی
اسکلت سلولی شبکه ای از رشته های مورد نیاز برای عملکرد مناسب از طیف وسیعی از فرایندهای سلولی از جمله تحرک سلول، تقسیم سلولی، قاچاق داخل سلولی و تعمیر و نگهداری از شکل سلول است. میکروتوبول ها، رشته های متوسط، و ریزرشته (همچنین به عنوان اسکلت سلولی اکتین شناخته می شود): اسکلت سلولی از سه سیستم مجزا رشته تشکیل شده است. این فعل و انفعالات پویا بین این رشته که با ساختارهای منحصر به فرد و توابع ارائه سلول است.
تعدادی از ساز و کارهای نظارتی، به کارگیری بسیاری از پروتئین اکتین اتصال، تکامل یافته اند برای کنترل دینامیک سیستم های اکتین رشته. اعتقاد بر این است که تروپومیوزین نقش محوری در این سیستم نظارتی بازی، موثر بر گروه رشته اکتین با سایرین.


